Динофлагелляты - одноклеточные протисты, которые внешне хорошо известны как аквариумистам, так и неспециалистам. Динофлагелляты, впервые обнаруженные Мюллером в 1773 году и позднее описанные Эренбергом в 1830-х годах при исследовании кремниевого известняка Мелового периода, предположительно возникли в Эдиакарскую эру или раньше, от 570 миллионов до миллиарда лет назад в период эволюции эукариот. Эти древние организмы прямо или косвенно ответственны за большую пользу и вред для человечества. Без динофлагеллятов около 7% населения земного шара лишилось бы пищи, некоторые страны перестали бы существовать и многие тысячи людей, ныне умерших, все еще были бы живы. Эти крошечные тельца также обеспечивают миллиарды долларов экономической выгоды. В этой статье я объясню, как такие, казалось бы, невероятные утверждения на самом деле верны.
Динофлагелляты, состоят примерно из 1200-2000 видов в 130 или более родах, и классифицируются по типу Dinoflagellata(Жгутиконосцев) и подразделению Pyrrophyta. Их название происходит от греческого слова dinos, что означает "кружение". Название дали характерные жгутики, которыми они обладают. Динофлагелляты, около половины из которых являются фотосинтетическими, а некоторые и биолюминесцентными, являются важными компонентами первичных продуктов океана. Они распространены во всех океанах и во всех океанических зонах и могут существовать в пелагических или бентических зонах или в тканях хозяина. Небольшой процент из них-пресноводные виды. Они, также, хорошо известны как паразиты или патогены, а некоторые даже являются симбионтами беспозвоночных. Динофлагелляты могут быть способны передвигаться и плавать (все живущие в водной среде) с помощью двух жгутиков. Эти жгутики, один расположенные вокруг клетки (поперечный жгутик), а другой направленный назад (продольный жгутик), являются диагностическими критериями этой группы.
Динофлагелляты обычно имеют внешнее покрытие, называемое тека, или амфиесма, из которого можно установить описательные признаки. По наличию или отсутствию хорошо развитых целлюлозосодержащих фекальных пластинок можно выделить две широкие группы динофлагеллятов, которые называются соответственно панцирными динофлагеллятами или беспанцирными (голыми) динофлагеллятами. Тека может быть разделена на две части поперечной канавкой, называемой пояском, или цингулюмом, которая окружает клетку. Часть теки над поясом - это эпитека, а часть ниже-гипотека. Пояс - это место расположения поперечного жгутика. Другая выемка, бороздка, проходит продольно и является местом прикрепления продольного жгутика. Передний конец клетки, Апекс, может иметь апикальный комплекс пор в панцирных динофлагеллятах, функция которых неизвестна. Задний конец клетки, антапекс, часто имеет выступающие шипы или выпуклости. Тека может быть сброшена, регенерирована или оставаться неразвитой в течение различных стадий жизни динофлагеллята.
Замечательные особенности динофлагеллятов, по-видимому, являются скорее правилом, чем исключением. Динофлагелляты могут испускать органеллы, похожие на нематоцисты кишечнополостных. Многие из них имеют многочисленные структуры вблизи периферии их внутренних клеток, называемые трихоцистами, черты, подобные тем, которые встречаются у инфузорий. Они выделяются при быстром увлажнении и, как полагают, функционируют при защите, выделении токсинов, секреции или захвате добычи. Другая особенность, обнаруженная у некоторых видов, - это мукоцисты, расположенные непосредственно под клеточной мембраной, которые выделяют слизистый материал, который может помочь клеткам прилипнуть к субстратам или может функционировать при захвате добычи. Третий тип эякулятивной органеллы встречается у некоторых видов. Это книдоцисты, и они очень похожи по строению на гидроидные нематоцисты. Они менее многочисленны, чем трихоцисты, и их функция неизвестна, но считается, что они участвуют в защите или захвате добычи. Другие необычные клеточные включения в динофлагеллятах могут включать плодоножку, наросты, глазные пятна, вирусоподобные частицы, параядерные тела и другие, и рассматриваются Spector (1994).
У динофлагеллятов есть и другие необычные особенности. Почти у каждого вида ядерные хромосомы постоянно конденсируются в состоянии, сходном с тем, которое обычно наблюдается до митоза в ядрах клеток других эукариот. Кроме того, динофлагеллятные ядра содержат во много раз больше ДНК, чем в большинстве других эукариотических клеток, что более подробно обсуждается ниже. По крайней мере, некоторые виды, такие как, полиплоидны. Динофлагелляты могут размножаться бесполым или половым путем. В первом случае клетка делится продольно путем деления, сохраняя гаплоидное состояние. Каждая гаплоидная часть может, выступать в качестве гамет. Жгутиковые гаплоидные клетки могут латерально сливаться и образовывать диплоидную зиготу, которая может оставаться подвижной. Однако при неблагоприятных условиях некоторые динофлагеллятные зиготы увеличиваются и утолщаются (стадия гипнозиготы), теряют подвижность и производят красные тела внутри клетки. Эта диплоидная форма, называемая цистой (или диноцистой), оседает на дно водной толщи и является "стадией покоя" в жизненном цикле динофлагеллята. Диноциста может иметь жесткое покрытие из вещества, называемого спорополленином, или она может быть внедрена с карбонатом кальция или кремнием. Диноцисты появляются, или прорастают, в подвижной стадии при благоприятных условиях окружающей среды, с другим витком деления, на этот раз мейозом, снова приводящим к гаплоидной клетке. Эта часть жизненного цикла называется гаплонтической.
Некоторые интерактивные и основные описания митоза и мейоза можно найти здесь
Some interactive and basic descriptions of mitosis and meiosis can be found here:
http://www.biologyinmotion.com/cell_division/http://www.pbs.org/wgbh/nova/miracle/divide.html#Трофическая стратегияМногие динофлагелляты являются фотосинтетическими и входят в число основных первичных продуцентов фитопланктона наряду с диатомовыми водорослями. Эта форма получения энергии позволяет считать около 50% динофлагеллятов автотрофными, хотя все, кроме нескольких видов, являются ауксотрофными для витамина В12, тиамина и биотина (обзор в Provasoli and Carlucci, 1974). Динофлагелляты обычно описываются как растения С3, хотя некоторые виды могут напоминать растения С4 при определенных условиях, и динофлагелляты, в целом, могут проявлять некоторые характеристики обоих типов. Разница между этими типами заключается в том, есть ли или три или четыре углеродных сахара и ферменты, используемые для фиксации CO2.
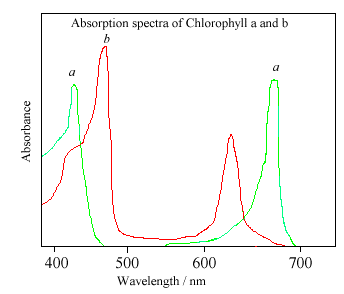
Хлоропласты - это мембраносвязанные органеллы, содержащиеся в фотосинтетических организмах, которые являются основными центрами сбора света и фотосинтеза и содержат большую часть фотосинтетических пигментов. Хлоропласты, обнаруженные в красных и зеленых водорослях, как известно, развились из симбиоза между цианобактерией и эукариотической клеткой более миллиарда лет назад. Основными светопоглощающими пигментами в большинстве хлоропластов растений являются хлорофиллы. инофлагелляты содержат как хлорофилл а (chl-a), так и хлорофилл с (chl-c), в то время как большинство растений и зеленых водорослей содержат в основном хлорофилл а и в меньшей степени хлорофилл b (chl-b). Хлорофиллы d и e также существуют в водорослях, причем первые в основном в некоторых красных водорослях. В то время как некоторые другие организмы помимо динофлагеллятов содержат chl-c, этот пигмент предполагает большее эволюционное неравенство между динофлагеллятами и большинством других "фитопланктонов". Это также дает преимущество в том, что фотосинтетические организмы, содержащие множество хлорофиллов, способны эффективно собирать световую энергию из более широкого диапазона длин волн света. В случае chl-b, более распространенного в зеленых водорослях, спектр смещается в сторону более длинных длин волн в зеленый спектр. Chl-c не имеет такого же большого пика в красном спектре, как chl-a, и можно предположить, что наличие chl-b было бы более выгодно для динофлагеллятов, поскольку меньшая конкуренция за свет является основной причиной наличия различных пигментов. Однако именно соединение хлорофиллов с перидинином, пигментом широкого спектра поглощения света, дает динофлагеллятам явное преимущество перед другими фитопланктонами. Хлорофиллы - это пигменты, в значительной степени ответственные за зеленую окраску растений. Первичные пики поглощения находятся на 430 Нм и 663 Нм, а также 434 Нм и 666 нм для chl-a и chl-c соответственно, что соответствует синей и красной областям спектра. Поскольку динофлагеллятные хлоропласты необычно содержат три мембраны, в отличие от нормальной одной или двух, считается, что они, вероятно, развили третичный эндосимбиоз с плазмидой, которая содержит дополнительный фотосинтетический пигментный комплекс перидинина (Morden and Sherwood 2002). Оранжево-красный пигмент перидинина поглощает очень широко, с максимумом на около 480нм и другим небольшим всплеском на 520нм. Объединенные единицы каротиноид-хлорофилл-белковых комплексов (ПХФ-комплекс), состоящие в основном из перидинина, хлорофилла а и одного из 12-20 белков, образуют множественные комплексы, где, что интересно, взаимодействие хлорофилла с белком перидинина сдвигает пики поглощения chl-a вверх примерно на 10 нм.
Один из спектров фотосентических пигментов:
 | Вот изображение какого-то Noctiluca, вероятно N. miliaris. Предположительно, это самые крупные динофлагелляты. Им не хватает хлорофилла, и они едят более мелких простейших. Они также блестяще биолюминесцентны, на что указывает их название, поскольку "noctiluca" означает "ночной свет".- Они имеют форму кувшинки, и то, что кажется "нитями" на этом изображении, - это большие жгутики, которые используются для поимки добычи. Фото и подпись: Рональд Шимек.
Другие пигменты используются косвенно в качестве вспомогательных пигментов. Некоторые из них обладают окислительными способностями и используются в цепях переноса электронов, которые являются частью как светлых, так и темных реакций фотосинтеза. Некоторые даже имеют дублирующие функции, добавляя другие уровни функции к фотосинтетической клетке. Например, фукоксантин может присутствовать в качестве вспомогательного пигмента у перидининсодержащих видов, в то время как у некоторых других он может заменить перидинин. Фукоксантин является распространенным каротиноидом в первую очередь у диатомовых и динофлагеллятов. |
Каротиноиды - это вспомогательные пигменты, которые отвечают за преимущественно желтую и оранжевую окраску и поглощают свет с длиной волны преимущественно между 450 Нм и 550 Нм. Их цвет обычно маскируется присутствием хлорофилла, но в динофлагеллятах хлорофиллы играют вторую скрипку перед перидинином. Каротиноиды состоят из каротинов и каротиноидов (ксантофиллов). Каротины обладают многочисленными вторичными функциями, но могут быть наиболее важными для зооксантелл, действуя как антиоксиданты. Ксантофиллы состоят из оксигенированных каротинов, таких как неоксантин, виолаксантин и лютеин, которые обеспечивают характерную окраску путем абсорбции, либо функционально, либо случайно. Фукоксантин представляет собой желто-зеленый пигмент с первичным поглощением около 530 Нм, что характерно для динофлагеллятов. Также важное значение в динофлагеллятах имеют ксантофиллы диноксантин и диадиноксантин, которые играют роль в предотвращении фотоокислительного повреждения фотосинтетического аппарата. Необычным для эукариот является также то, что динофлагелляты демонстрируют отчетливые циркадные ритмы, в первую очередь за счет суточной миграции хлоропластов внутри клеток.
| Цвет: | Название: | Пик поглощения (нм): |
| оранжевый | бета-каротин | 447, 449 |
| сине-зеленый | *хролофилла | 662, 429 |
| оранжево-желтый | неизвестно | |
| желтый | *диадиноксантин | 476, 446 |
| оранжево-красный | *перидинин (64%) | 480 |
| светло зеленый | хролофилл c1, c2, c3 | 461, 583, 644 |
| розовый | неизвестно | 464 |
| yellow | *диноксантин | 470, 440 |
| *диатоксантин | 427, 454, 482 |
| кирпичный | нео-перидинин | |
| желто-зеленый | фукоксантин | 500 |
| зеаксантин | 455 |
| P-457 (производное неоксантина) | 457 |
*основные компонентыТаблица 1. Пигменты, обнаруженные в зооксантеллах кораллов, содержат многочисленные виды динофлагеллятов.Возможно, наибольший интерес для аквариумистов представляют беспанцирные морские динофлагелляты многих видов - морские симбионты, известные как зооксантеллы, которые обитают в гастродермисе большинства герматипических (риф-строительных) коралловых полипов. Однако динофлагелляты играют сходную симбиотическую роль с другими морскими беспозвоночными, включая морские анемоны, радиолярии, губки, фораминифераны, турбеллярии, медузы, моллюски и другие группы. Большая часть золотистого или коричневого цвета кораллов обусловлена зооксантеллами, и в частности содержанием в их составе ксантофилла. Степень, в которой этот цвет влияет на общий цвет кораллов, зависит от многих факторов, включая генетику (наследуемый фенотип), плотность пигмента, плотность клеток водорослей и формированием флуоресцирующих белков животными. Плотность водорослевых клеток и пигментов может быть функцией света (более яркий свет производит более низкую плотность пигмента/клеток, а менее яркий свет производит более темно окрашенные колонии из более высокой плотности динофлагеллятных клеток/концентрации пигмента) и питательных веществ (больше азота вызывает более высокую плотность динофлагеллятных клеток). Тем не менее симбиоз с кораллами позволил коралловым рифам развиваться, поскольку партнерство позволяет скорости кальцификации кораллов опережать естественные деградационные и эродирующие процессы. Ускоряя развитие кораллов и рост коралловых рифов, динофлагелляты вносят большой вклад в прибрежный буфер многих стран, среду обитания, которая позволяет вести продуктивный промысел, обеспечивающий до 7% мирового потребления белка, и косвенно ответственны за миллиарды долларов доходов от туризма и рекреационного использования, которые сосредоточены на деятельности коралловых рифов.
 Маленькие, круглые, беспацирные динофлагелляты, лишенные жгутиков, являются эндосимбионтами кораллов и видны здесь как почти полный монослой в гастродермисе Acropora cervicornis (верхний клеточный слой).
Маленькие, круглые, беспацирные динофлагелляты, лишенные жгутиков, являются эндосимбионтами кораллов и видны здесь как почти полный монослой в гастродермисе Acropora cervicornis (верхний клеточный слой).
Фото Eric Borneman.Однако не все динофлагелляты автотрофны, а некоторые вообще не фотосинтезируют. Они также могут существовать по нескольким различным гетеротрофным стратегиям, включая виды, которые являются фаготрофными (поглощающими целые клетки), сапрофитными (питающимися разлагающимся веществом), паразитическими (питающимися непосредственно другими организмами) и мутуалистическими (живущими во взаимовыгодных симбиозах). Травоядность возможна у многих видов, а некоторые виды обладают мощными целлюлолитическими ферментами для разрушения клеточных стенок растений. Фаготрофные виды могут прикрепляться к поверхности своей добычи и затем развивать ризоподии, которые обволакивают клетку. Известно, что некоторые динофлагелляты питаются яйцами (особенно яйцами копепод), одноклеточными и нитевидными водорослями, бактериями и другими микроорганизмами.
Токсичные Динофлагелляты Возможно, самым печально известным аспектом динофлагеллятов является способность некоторых видов производить токсины. Цветение производящих токсин динофлагеллятов называют "красными приливами", и эти часто сезонные события могут быть неожиданными. Динофлагелляты также ответственны за
сигуатеру и другие отравления моллюсков. Вредные цветение динофлагеллятов, производимое 20-70 токсичными видами, очень похожи на цветение нетоксичных видов и, вероятно, происходят путем конкурентного исключения. Однако все токсичные виды являются фотосинтетическими, существуют в эстуариях или неритах (прибрежные воды, покрывающие континентальный шельф) и производят водорастворимые или липидорастворимые токсины.
"Красный прилив", более правильно называть вредным цветением водорослей, является несколько неправильным названием, поскольку вода редко становится красной, но чаще оранжевой, что является результатом быстрого цветения и плотной популяции (до 100 миллионов клеток/л) одного или нескольких видов динофлагеллятов (или диатомовых водорослей). Они могут быть проблематичными, поскольку некоторые из быстро цветущих видов производят мощные нейротоксины, называемые сакситоксинами (и связанные с ними токсины). Эти токсины накапливаются в суспензионных кормушках, таких как съедобные мидии и моллюски, а также могут накапливаться или убивать рыбу или других животных, включая человека, которые едят этих моллюсков. Появление симптомов, называемых паралитическим отравлением моллюсками, может произойти почти сразу после приема пищи. При вдыхании смерть может наступить в течение нескольких минут. По иронии судьбы, сакситоксин и тетрадотоксин в ничтожных количествах являются частью "опасного удовольствия" от употребления фугу, сашими из различных иглобрюхих. Оба действуют сходным образом, блокируя натриевые каналы. Находясь в прошлом году в Фукуоке, возможно, основном центре, где употребляют фугу в Японии, я решил не искушать судьбу на прием сакситоксина/тетратодотоксина. Другие динофлагеллятные виды могут вырабатывать бреветоксины, вызывающие симптомы, сходные с симптомами нейротоксического отравления моллюсками. Массовая гибель рыбы вдоль побережья Мексиканского залива была вызвана производящими бреветоксин динофлагеллятными видами, и аэрозолизация морской пыли во время красных приливов может вызвать болезни у тех, кто живет на побережье или посещает его. Наиболее распространенным заболеванием морскими токсинами является сигуатера, неврологическое расстройство желудочно-кишечного тракта и сердечно-сосудистой системы, вызванное накоплением производимого динофлагеллятами сигуатоксина в зараженной рыбе. Чем выше трофический уровень рыбы, тем более концентрирован сигуатоксин, а барракуды, угри, морские окуни, щелкуны и домкраты печально известны тем, что вызывают случаи сигуатоксикоза у людей после употребления в пищу. Для получения дополнительной информации о Ciguatera, перейдите по этой ссылке:
http://www.cdc.gov/nceh/ciguatera/default.htm Барракуда, хотя и вкусная, является одной из наиболее вероятных причин отравления сигуатерой, если ее съесть, особенно более старую и крупную рыбу.
Барракуда, хотя и вкусная, является одной из наиболее вероятных причин отравления сигуатерой, если ее съесть, особенно более старую и крупную рыбу.В результате исследований в 1988 году были найдены токсичные динофлагелляты
Pfiesteria piscicida. Этот вид был ответственен за массовые убийства рыб вдоль Атлантического побережья Соединенных Штатов и производит по крайней мере один очень мощный нейротоксин. Цветение, происходящее главным образом в эстуарийных местообитаниях, недолговечно, часто длится всего несколько часов. Что делает Фистерии настолько интригующими, так это то, что в ее жизненном цикле есть по крайней мере 24 формы, включая нормальные нетоксичные формы фотосинтеза, токсичные и нетоксичные формы кисты и хищную форму. Один токсин, вырабатываемый на стадии кисты, оглушает рыбу, облегчая прикрепление динофлагеллята. Тот же токсин вызывает некроз тканей и открытые язвы. Еще один токсин влияет на дыхание рыб. Затем динофлагеллят принимает микропредуктивную роль и питается растворяющейся тканью. Для получения дополнительной информации посетите:
http://www.epa.gov...pfiesteria/fact.html.
Динофлагелляты в Аквариуме: "Сопливые" Динофлагелляты и Паразиты РыбПомимо мер предосторожности, которые должны быть приняты в отношении токсичных динофлагеллятов, которые, безусловно, потенциально присутствуют в морских аквариумах, и очевидного интереса к симбиотическим видам, таким как зооксантеллы, аквариумисты, хорошо знакомы с историями о коричневых, слизистых, сопливых водорослях, которые поглощают пузырьки воздуха и покрывают поверхности живых камней, стенки аквариумов и даже кораллы. Такие периодические цветения часто отмечаются весной и, по-видимому, коррелируют с гибелью рыб и особенно с летаргией или параличом и гибелью травоядных улиток. Они широко освещены у Sprung и Delbeek (1994), наряду с методами искоренения. Такие случаи действительно могут быть цветением токсичных динофлагеллятов. Однако есть и другие токсичные микроводоросли, другие слизистые сопливые водоросли и другие водоросли, которые поглощают пузырьки воздуха. Пузырьки воздуха, вероятно, являются кислородом, вырабатываемым фотосинтезом, и из многих матирующих или пленкообразующих микроорганизмов фотосинтетические протисты, водоросли и цианобактерии могут выглядеть очень похожими друг на друга. Цианобактерии, в частности, также хорошо известны тем, что производят токсины.
Во время погружений как в Карибском бассейне, так и в Тихом океане я заметил сопливый коричневый материал с пузырьками газа, который, по-видимому, является локализованными областями того, что описывается как динофлагелляты в рифовых аквариумах. Недавно было обнаружено, что эти скопления на рифах состоят из хризофитов, в частности Chrysocystis fragilis (Schaffelke et al. 2004). Интересно, что многие хризофиты имеют несколько жизненных стадий, могут вырабатывать токсины, особенно во время цветения (Boenig and Stadler 2004), содержат пигменты, подобные тем, которые содержатся в динофлагеллятах, и могут иметь два жгутика. Они золотисто-коричневого цвета, и их легко спутать с динофлагеллятами, даже если исследовать их под микроскопом. Они также коррелируют со смертностью тканей и признаками обесцвечивания, когда они покрывают живые кораллы в дикой природе, подобно тому, что наблюдается в аквариумах. Более детально о хризофитах можно почитать здесь:
http://www.ucmp.berkeley.edu/chromista/chrysophyta.html.
 Хризофиты на рифовой скале между Millepeora alcicornis и Montastraea faveolata. Фото Эрика Борнемана.
Хризофиты на рифовой скале между Millepeora alcicornis и Montastraea faveolata. Фото Эрика Борнемана.
Последние и, пожалуй, самые известные группы динофлагеллятов являются паразитами рыб, Amyloodinium ocellatum и Cryptocaryon irritans, более известные как морской бархат и Ихтиофтириоз, соответственно. Эти распространенные и вызывающие беспокойство паразиты широко освещены в аквариумной и научной литературе.ЗаключениеДинофлагелляты эволюционирующий древний вид, включающий чрезвычайно разнообразные одноклеточные организмы. Используя миксотрофные стратегии и обладая многочисленными редкими и характерными чертами, динофлагелляты хорошо приспособлены к выживанию. Фотосинтетические виды имеют хлоропласты и митохондрии, которые, как полагают, первоначально представляют собой эндосимбиозы с примитивной прокариотической клеткой. Динофлагелляты с тройной мембраной, заключенной в пердининсодержащий хлоропласт, и склонностью к образованию паразитических или мутуалистических симбиозов с другими морскими видами, вероятно, представляют собой благоприятно адаптивный образ жизни. Понимание природы их экологии и биологии лучше вооружает аквариумистов, чтобы иметь дело как с неприятными, так и с полезными видами. По крайней мере, коричневые кораллы могут приобрести совершенно новый "свет" в глазах своих созерцателей.
Оригинал статьи

